
El funcionamiento de los ecosistemas influye en la aptitud de las especies en super predadores
El cambio global está afectando gravemente el funcionamiento de los ecosistemas y la biodiversidad a nivel mundial. Los atributos funcionales de los ecosistemas (AFE) detectados remotamente son descriptores integradores del cambio ambiental y están estrechamente relacionados con los procesos que afectan directamente las cadenas alimentarias a través de las cascadas tróficas. Este estudio realizado en Galicia y puso él foco en el águila real, evalúa el éxito reproductivo en esta especie un depredador ápice en el nivel trófico superior, durante un período de 17 años a lo largo de un gradiente bioclimático
Biodiversidad27/04/2022
El cambio global antropogénico está afectando gravemente el funcionamiento de los ecosistemas y la biodiversidad a nivel mundial (Flombaum et al. 2017; Rillig et al. 2019; Díaz et al. 2020). Un creciente cuerpo de trabajo ha demostrado cómo el calentamiento climático, el cambio en el uso de la tierra y sus efectos conjuntos han estado, directa o indirectamente, afectando la biodiversidad y el funcionamiento de los ecosistemas, con efectos secundarios difíciles de predecir en sus relaciones complejas y dinámicas (Carnicer et al 2011; Dib et al. 2020). Una mejor comprensión de las relaciones entre el funcionamiento de los ecosistemas y la biodiversidad en los diferentes niveles tróficos es esencial para ayudar a los administradores a responder adecuadamente a las amenazas que el cambio global representa para la biodiversidad y la salud de los ecosistemas (Thébault y Loreau 2006). También es fundamental desarrollar sistemas rentables de monitoreo y alerta temprana para responder proactivamente a estas amenazas con estrategias de gestión efectivas (Cabello et al. 2012; Anderson 2018; Aragón et al. 2019).
Los super depredadores se encuentran en la parte superior de la cadena alimentaria y desempeñan un papel fundamental en la dinámica de la población de los niveles tróficos inferiores (Wallach et al. 2015). Los efectos interactivos del calentamiento global del clima, los cambios regionales en el uso de la tierra y los regímenes alterados de perturbación pueden afectar de manera crítica la dinámica de la población de estas especies clave, con impactos posteriores en otros niveles tróficos a través de interacciones depredador-presa (Guiden et al., 2019). Hasta ahora, la mayoría de los estudios ecológicos sobre los superdepredadores se basan en enfoques de modelado respaldados por datos macroclimáticos (obtenidos de interpolaciones de estaciones meteorológicas) y características estáticas del hábitat (ver, por ejemplo, López-López et al. 2007; Tapia et al. 2007 ). A pesar de la creciente disponibilidad y calidad de los conjuntos de datos macroclimáticos, estos datos no pueden representar con precisión las condiciones microclimáticas a menudo impulsadas por los patrones topográficos y de vegetación local, y no logran capturar las características críticas de su nicho ecológico (Amiri et al. 2020). Estas limitaciones resaltan la necesidad de utilizar descriptores mas proximales de las condiciones de hábitat ‘reales’—en el sentido más amplio—que la especie está experimentando a nivel del suelo.

Es ampliamente aceptado que el balance energético y la productividad primaria neta afectan la riqueza
de especies locales porque cuanta más energía hay disponible, más biomasa por unidad de área se puede sustentar (Phillips et al. 2008; Brasil et al. 2019). Más biomasa permite que más individuos coexistan en un área, lo que potencialmente resulta en más energía disponible para la reproducción, lo que reduce las tasas de extinción (Connell y Orians 1964). Por lo tanto, la no inclusión de procesos ecosistémicos críticos cómo la productividad primaria o el balance energético superficial puede socavar nuestra capacidad para monitorear estas especies clave. Por ejemplo, recientemente se descubrió que la variabilidad temporal en la productividad primaria afecta las fluctuaciones de la calidad del hábitat y la dinámica de la metapoblación de las especies consumidoras al afectar los niveles de energía disponibles como recursos alimentarios (Fernández et al. 2016; Letnic et al. 2018). La fenología del hielo marino y los pulsos de productividad primaria dan forma al éxito reproductivo de las aves marinas del Ártico (Ramírez et al. 2017).
La disciplina de la teledetección ha evolucionado en gran medida durante las últimas décadas y actualmente permite monitorear y evaluar los componentes clave del funcionamiento de los ecosistemas (es decir, el flujo biogeoquímico de energía y materia dentro de los ecosistemas) (Lovett et al. 2006; Jax 2010; Van den Broeke et al. 2011; Lausch et al. 2016). A pesar de la creciente disponibilidad de descriptores de funciones ecosistémicas obtenidos mediante sensores remotos, su contribución potencial al monitoreo de especies en los niveles tróficos superiores sigue siendo poco estudiada. Entre otros, la temperatura de la superficie terrestre y el albedo, y estacional la productividad y la fenología son buenos descriptores de las funciones de los ecosistemas, siendo relativamente fáciles de obtener a partir de imágenes de teledetección (Metz et al. 2014; Zhao et al. 2018b; Aragón et al. 2019).
La temperatura de la superficie terrestre (LST) es un descriptor clave del balance de energía radiativa de la superficie de la Tierra (Hulley et al. 2019). La variación intra e interanual de este descriptor se usa a menudo como proxy de la dinámica del calor sensible (Li et al. 2013; Metz et al. 2014). Típicamente, la LST derivada de satélite se recupera estimando la radiancia emitida desde la superficie que se obtiene al corregir atmosféricamente la radiancia del sensor. LST mide la emisión de radiación térmica de la superficie terrestre donde la energía solar entrante interactúa y calienta el suelo, o la superficie del dosel en áreas con vegetación (Oyler et al. 2019). Por lo tanto, LST es un buen indicador de la partición de energía en el límite entre la superficie terrestre y la atmósfera y es especialmente sensible a las variaciones en el tipo de cobertura terrestre, la densidad de la vegetación o la humedad del suelo (Hulley et al. 2019). De hecho, la LST está determinada en gran medida por los gradientes climáticos y los patrones de productividad de la vegetación, y también está influenciada localmente por las condiciones topográficas y microclimáticas (Oyler et al. 2019). Además, las perturbaciones tanto naturales como antropogénicas (por ejemplo, incendios forestales) pueden ser otra fuente importante de variación espaciotemporal debido a cambios abruptos en los patrones de LST.

El albedo, definido cómo la radiación solar reflejada en la radiación solar entrante, es otro componente clave del funcionamiento del ecosistema relacionado con el balance de energía superficial. Puede medirse desde el espacio integrando información recuperada de sensores multibanda específicos basados en las propiedades de anisotropía definidas por funciones de distribución de reflectancia bidireccional. El albedo varía en grandes áreas debido a las variaciones en las propiedades de la superficie terrestre, como el tipo de cobertura terrestre, la cobertura de vegetación verde, la rugosidad de la superficie o la humedad del suelo. Por lo tanto, puede verse afectado por cambios en el uso de la tierra inducidos por el hombre y circuitos de retroalimentación del cambio climático (Hu et al., 2019). Además, la perturbación del fuego puede cambiar abruptamente la superficie terrestre con efectos duraderos en el albedo (Dintwe et al. 2017).
Este trabajo tiene como objetivo mejorar nuestra comprensión sobre cómo el funcionamiento de los ecosistemas influye en las especies clave en los niveles tróficos superiores aprovechando el gran potencial de la teledetección para rastrear los flujos de materia y energía desde el espacio. Esperamos que los atributos funcionales del ecosistema detectados remotamente (derivados de productos de detección remota como LST, Albedo y productividad de la vegetación) puedan, al menos, explicar parcialmente la aptitud ecológica de un superdepredador: el águila real. Para hacerlo, aprovechamos una base de datos de series temporales a largo plazo del éxito reproductivo del águila real durante un período de 17 años a través de un gradiente bioclimático. Computamos una base de datos integral de atributos funcionales del ecosistema a partir de tres productos satelitales MODIS (espectrorradiómetro de imágenes de resolución moderada) relacionados con el ciclo del carbono, la dinámica del calor y el balance radiativo. También evaluamos el posible retraso en la respuesta del águila real al fuego, un disruptor crítico del presupuesto de energía de superficie en nuestra región. El área de estudio es Galicia, región situada en el Noroeste de España (c. 29.575 km2 ) (Figura 1).
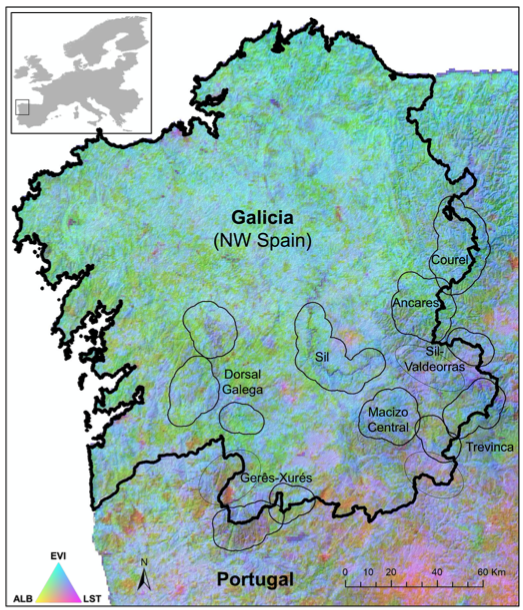
Figura 1. Ubicación del área de estudio en Europa. Imagen de valor de saturación de tono (espacio de color HSV) calculada a partir de la combinación de productos MODIS que representan la fenología (fecha del albedo mínimo anual (ALB), en el componente circular de tono), estacionalidad o variación intraanual (desviación estándar de la temperatura de la superficie terrestre (LST), componente de saturación), y productividad primaria (promedio anual del Índice de Vegetación Mejorado (EVI), componente de valor). Los polígonos representan las áreas montañosas donde se ubican los territorios de reproducción históricos y actuales.
El clima es de transición entre atlántico y mediterráneo (Rodríguez Guitian y Ramil-Rego 2007; Rodríguez-Lado et al. 2016). Promedio la precipitación anual oscila entre 700 y 2000 mm, y la temperatura media anual entre 8,5 y 15 C (Rodríguez- Lado et al. 2016). Las altitudes son máximas (hasta 2071 msnm) en las montañas orientales y mínimas en la costa y valles fluviales. Las superficies forestales cubren más del 70% de la región de estudio, siendo actualmente una de las zonas más densamente arboladas de España. En las zonas de montaña, los sistemas agropastoriles tradicionales extensivos se han conservado hasta la actualidad, aunque han sido abandonados paulatinamente desde mediados del siglo XX, siendo recolonizados por bosques naturales y plantaciones de árboles de rápido crecimiento (Regos et al. 2015; Salaverri et al. 2018). El área de estudio está relativamente escasamente poblada, con algunas áreas grandes que tienen menos de 10 habitantes/km2 en las áreas montañosas del este. El régimen de incendios provocado por el hombre (incluidas las causas de negligencia e incendio provocado) expone el área de estudio a una frecuencia inusualmente alta de incendios forestales (Chas-Amil et al. 2010). Entre 1968 y 2016 se produjeron más de 256.000 incendios en Galicia, quemando más de 1.931.000 ha (ca el 63% del territorio) (Regos 2018).
El águila real es un super depredador, situado en el nivel superior de la cascada trófica. Esta rapaz puede describirse como depredador tanto generalista como oportunista, y se alimenta de una amplia variedad de especies en todo su rango de distribución mundial (Watson 2010; Bedrosian et al. 2017). Típicamente, esta especie se alimenta de mamíferos de tamaño mediano que oscilan entre 0,5 y 4,0 kg, aunque amplía su dieta cuando las especies de presa preferidas son escasas (Steenhof y Kochert 1988; Watson 2010).
Un depredador ápice como el águila real tiene altos requerimientos de alimentos y energía, por lo que su rendimiento y aptitud ecológica, medidos por el éxito reproductivo, depende en gran medida de las fluctuaciones interanuales de la disponibilidad de presas, en su mayoría consumidores primarios, que a su vez dependen del balance energético de la superficie. dinámica y funcionalidad de los ecosistemas (Fernández et al. 2016; Preston et al. 2017). El área de estudio constituye el límite noroeste del área de distribución del águila real en la Península Ibérica, donde la especie se considera actualmente ‘Casi Amenazada’ (Madroño et al. 2005). En Galicia, el águila real tiene una población de 14 parejas reproductoras confirmadas, con otras 9 parejas compartidas con regiones vecinas del norte de Portugal y España (Gil-Carrera et al. 2016). Las áreas aptas para la anidación en el sitio de estudio se caracterizan por estar ubicadas en sierras entre 255 y 1443 msnm, cañones de ríos de fuertes pendientes, influencia climática mediterránea y zonas de baja presión humana (Lado y Tapia 2012). Los principales problemas de conservación están asociados con las limitaciones del hábitat, como el suministro inadecuado de alimentos, la escasa disponibilidad de sitios de anidación, la perturbación humana en las áreas de reproducción, como la electrocución y los cambios en el uso del suelo (Madroño et al. 2005; Tapia et al. 2007). Además, esta pequeña población habita una zona bioclimática de transición relativamente aislada de otros núcleos de la Península Ibérica (Tapia et al. 2007; Lado y Tapia 2012), y un aumento de la mortalidad puede tener un impacto importante en sus parámetros demográficos (Whitfield et al. 2004).
Para la realización de este trabajo, se han realizado campañas de reconocimiento de campo anuales (búsqueda a pie y en la carretera desde un vehículo 4 9 4) entre 2001 y 2017 en áreas aptas para la anidación con el fin de monitorear los territorios de reproducción conocidos y detectar nuevos (ver (Tapia et al. 2007, 2009; Gil y Tapia 2009) para detalles sobre el estudio de campo e inventario de los antiguos territorios de reproducción). La distribución del águila real en el área de estudio es bien conocida, siendo la tasa de pseudo-ausencias baja (N = 397 observaciones). El conjunto de datos final incluye 59 territorios de cría históricos y actualmente ocupados en Galicia y las regiones limítrofes (hasta 10 km de la frontera con Galicia). Estos 59 territorios de reproducción están ubicados en diferentes sistemas montañosos con condiciones geomorfológicas y bioclimáticas particulares (Figura 1). Caracterizamos el ámbito de hogar de cada pareja reproductora considerando radios de 5 km desde el nido (es decir, incluyendo el sitio del nido, las áreas de post-emplumamiento y alimentación; Tapia y Zuberogoitia 2018). Durante el período de 17 años, se registraron 168 parejas reproductoras en los 59 territorios, aunque solo se confirmaron 81. Algunas parejas reproductoras han cambiado sus nidos durante el período de estudio dentro del mismo territorio de reproducción. En estos casos, seleccionamos el punto central de ubicación del nido en cada territorio para el análisis.

Junto con la inmigración y la emigración, la natalidad y la mortalidad son los parámetros demográficos más importantes que determinan las tendencias de año a año en las poblaciones locales de rapaces (Steenhof y Newton 2007). En el presente estudio nos enfocamos en el éxito reproductivo para cada territorio y año de monitoreo. El éxito reproductivo se define como la proporción de parejas que anidan y ponen que crían crías hasta la edad de emplumar. Consideramos que las parejas tienen éxito cuando se observan crías bien desarrolladas en el nido en algún momento antes de emplumar. Los estudios que consideran que los nidos con crías de cualquier edad son exitosos tienden a sobreestimar el éxito del nido porque no consideran la mortalidad que puede ocurrir tarde en el período de crianza (Steenhof y Newton 2007).
Se seleccionaron tres productos satelitales MODIS (espectrorradiómetro de imágenes de resolución moderada) para describir tres dimensiones del funcionamiento del ecosistema: (1) índice de vegetación mejorado como sustituto de la dinámica del ciclo del carbono; (2) la temperatura de la superficie terrestre como sustituto de la dinámica del calor sensible; y (3) Albedo como sustituto del balance radiativo (Paruelo et al. 2001b; Cabello et al. 2012). Además, incluimos (4) Área quemada, como descriptor de la perturbación del fuego, que puede afectar dramáticamente estos tres descriptores biofísicos de las funciones del ecosistema (Bowman et al. 2009; Archibald et al. 2018). Los cuatro productos seleccionados por MODIS fueron:
- Índice de vegetación mejorado (EVI; MOD13Q1.006; 16 días L3 Global 250 m). El EVI es un descriptor de las ganancias de carbono, ya que se sabe que es más confiable en situaciones de cobertura vegetal alta y baja que el índice de vegetación de diferencia normalizada (NDVI), y resistente a las influencias del suelo, las señales de fondo del dosel y los efectos atmosféricos en el índice de vegetación. valores (Potter et al. 2007). Los valores de EVI oscilaron entre -1 y 1, y la vegetación saludable generalmente mantuvo valores entre 0,20 y 0,80.
- Temperatura de la superficie terrestre (LST; MOD11A2.006; 8 días L3 Global 1 km). LST es un buen descriptor del balance de energía en la superficie de la Tierra y uno de los parámetros clave en la física de los procesos de la superficie terrestre desde escalas regionales hasta globales. Además, LST está directamente relacionado con los regímenes ambientales primarios y con los atributos de calidad del hábitat (por ejemplo, productividad, estructura de la vegetación, tipo de cobertura del suelo; Cord y Rödder 2011). Las temperaturas (LST) oscilaron entre -25 C y 45 C.
- Albedo (ALB; MCD43A3.006; Daily L3 Global 500 m). ALB es un sustituto de las propiedades de la superficie, como la extensión y la naturaleza de la cubierta vegetal, y se ve afectado por el cambio de los factores biofísicos de la superficie terrestre, como la vegetación, la temperatura de la superficie terrestre y la humedad del suelo (Zhao et al. 2018a). Los valores de ALB oscilaron entre 0 y 1 (la nieve fresca y el suelo desnudo suelen caer alrededor de 0,9).
- Área Quemada (BA; colección 6 producto MODIS Área Quemada). BA es un buen descriptor de las interrupciones del presupuesto de energía de la superficie, ya que los incendios forestales representan un intercambio crítico de energía y materia entre la superficie y la atmósfera a través de la combustión (Bowman et al. 2009; Archibald et al. 2018). Los valores de BA oscilaron entre 0 y 1 (valores hasta 1 para aquellas áreas totalmente afectadas por el fuego).
Todos los predictores MODIS nativos y derivados se calcularon para el período de tiempo 2001–2017 con una resolución de 1 km y luego se agregaron (valores medios) en el nivel de búfer de 5 km mediante el uso de la plataforma computacional Google Earth Engine basada en la nube (Gorelick y otros 2017). Para cada una de estas 3 dimensiones del funcionamiento del ecosistema (EVI, LST y ALB), derivamos las siguientes 7 métricas resumidas de su dinámica estacional: media anual (sustituto de la cantidad total anual); máximos y mínimos anuales (descriptores de los extremos anuales); desviación estándar estacional y coeficiente de variación (descriptores de estacionalidad); y fechas de máximo y mínimo (descriptores de fenología) (ver Tabla 1). El conjunto de datos completo incluía 21 descriptores del funcionamiento del ecosistema (7 métricas 9 3 dimensiones) como predictores candidatos en competencia. Cada uno de estos 21 atributos funcionales del ecosistema detectados remotamente se calcularon para cada año para medir la variabilidad interanual. Además, creamos variables que describen un lapso de tiempo en la respuesta del águila real al fuego. Para ello, estimamos el área quemada de cada año en cada zona de amortiguamiento de 5 km alrededor del nido. Luego, se crearon versiones retrasadas de la serie temporal cambiando el año en que ocurrió el evento de incendio (de 1 a 5 años después del evento de incendio).
Para evaluar el efecto del funcionamiento del ecosistema en el éxito reproductivo del águila real, realizamos modelos de exposición logística de supervivencia del nido (Brown et al. 2013) mediante el uso de modelos lineales generalizados de efectos mixtos (GLMM; Bolker et al. 2009) con paquetes R ‘lmer4’ (Bates et al. 2014).
Estos modelos de supervivencia del nido de exposición logística son similares a los modelos de regresión logística en que la variable de respuesta es binomial (éxito o fracaso reproductivo), pero la función de enlace se modifica del enlace logit para considerar los días de exposición del nido (Shaffer 2004)—101 días para el águila real (ver Brown et al. 2013 y referencias allí). Los GLMM se utilizaron para dar cuenta de la estructura jerárquica del conjunto de datos de especies, en el que cada conjunto de datos subyacente cubre una temporada de reproducción diferente (es decir, un año) en cada sistema montañoso (Bolker et al. 2009; Harrison et al. 2018). Incluimos los descriptores de detección remota de las funciones del ecosistema como factores fijos y ‘año’ y ‘sistema montañoso’ como efectos aleatorios anidados. Sobre la base de esta estructura de efectos aleatorios, el componente de efectos fijos se modificó mediante la función ‘dredge’ disponible en el paquete R ‘MuMIn’ para ejecutar la selección de modelos automatizada con subconjuntos de todas las combinaciones (válidas) de variables explicativas (Barton 2016). Calculamos cada modelo delta AIC (DAIC) para verificar la fuerza de la evidencia a favor de cada modelo. Los valores de Delta AIC por debajo de 4 sugieren evidencia sustancial para el modelo (Burnham y Anderson 2002; Arnold 2010). Se calculó un modelo final promedio a partir de aquellos con DAIC por debajo de 4 mediante el uso de la función ‘model.avg’ disponible en el paquete R ‘MuMIn’. La importancia de cada predictor se obtuvo sumando los pesos de Akaike (Wi) a los modelos en los que está presente esa variable (Burnham y Anderson 2002; Arnold 2010).
Para evitar incluir variables altamente correlacionadas en ajuste del modelo, eliminamos las variables de detección remota con los coeficientes de correlación de Pearson anteriores 0,7 o inferior – 0,7 y factor de inflación variable mayor que 3. Por lo tanto, a partir del conjunto de datos inicial de 21 Atributos funcionales del ecosistema detectados remotamente, solo retuvimos 12 variables no correlacionadas relacionadas con
el ciclo del carbono (4 variables del tiempo EVI serie), dinámica del calor sensible (4 variables de
serie temporal LST) y balance de radiancia (4 variables de la serie de tiempo de Albedo).
Los resultados se obtienen en base al análisis del éxito reproductor del águila real durante 17 años en Galicia. Los modelos que presentaban mayor serán aquellos que incluían variables relacionadas con el ciclo de carbono en el ecosistema, el cual está relacionado con la productividad vegetal. Si los modelos se obtienen incluyendo variables funcionales del ecosistema, en especial las variables relacionadas con el balance energético superficial), también presentan una buena respuesta y predicción, algo inferior que los modelos basados en la productividad vegetal. El modelo integrador (incluyendo todas las variables anteriores), muestra un buen rendimiento. Si se incluyen variables como la alteración del hábitat, más concretamente el efecto de los incendios transcurridos 3 años, se incrementaría el éxito en las predicciones de los modelos.
Nuestro estudio proporciona evidencia de la influencia de el ecosistema funcionando en la aptitud de un vértice depredador (es decir, un depredador en el nivel trófico superior). Desde el conjunto de datos inicial de 12 de forma remota descriptores funcionales del ecosistema detectados, 4 de ellos (la desviación estándar y la media anual de LST, la media anual de EVI y la fecha del mínimo Albedo) afectaron el desempeño reproductivo de esta población residual de Águila Real en el NW de Iberia en los últimos 17 años. Perturbación de fuego también afectó la aptitud ecológica de este depredador ápice, pero con un efecto positivo limitado a los 3 años después del incendio (es decir, una respuesta retardada en el tiempo a la energía superficial interrupciones presupuestarias). A pesar de la moderada capacidad predictiva de nuestros modelos (valores AUC desde 0,71 a 0,8), debemos ser conscientes de la conocida relevancia de otros factores abióticos y bióticos, que no se incluyen aquí—que determinan el éxito reproductivo de este super depredador (como la presión humana o los brotes de la enfermedad de los conejos).
La desviación estándar del LST intraanual dinámica—un descriptor del calor estacional dinámica: afectó positivamente el sistema reproductivo comportamiento de la especie (ver Figura 2 y Tabla 2). De acuerdo con nuestros resultados, cuanto mayor sea la variación interanual en LST, mayor es la probabilidad de éxito reproductivo. Este patrón podría explicarse por el marcado gradiente bioclimático de nuestra área de estudio, en la transición entre las regiones Eurosiberiana y Mediterránea (Rodríguez Guitian y Ramil-Rego 2007). Los territorios de cría han sido tradicionalmente más productiva en la parte oriental de nuestra región (Gil y Tapia 2009; Lado y Tapia 2012), caracterizada por una mayor variación climática intraanual (es decir, veranos secos e inviernos templados y húmedos) que la zona atlántica eurosiberiana (caracterizada por un clima húmedo moderado por la influencia de el océano, con inviernos fríos y ausencia de una estación seca diferenciada). Esta correlación entre la variación estacional en LST y el reproductivo el rendimiento también se puede explicar por la altitud y gradientes topográficos, desde el Golden Eagle tiende a ocupar y reproducirse en áreas montañosas en salientes de acantilados con influencia climática mediterránea (Tapia et al. 2007; Lado y Tapia 2012), caracterizada por fuertes gradientes térmicos. En Además, una gran variación interanual en LST fue observado en los últimos 17 años en nuestra región, debido probablemente al efecto conjunto de fluctuaciones climáticas interanuales y disturbios por incendios (Goetz et al. 2007; Ueyama et al. 2014). Por lo tanto, las retroalimentaciones positivas entre el calentamiento climático y el aumento de la perturbación por incendios podrían favorecer el desempeño reproductivo de la especie en las próximas décadas en nuestra región, como se pronosticó anteriormente para otras águilas en España (Muñoz et al., 2013).
La media anual de LST fue otro descriptor relevante de la dinámica del calor estacional para explicar el éxito reproductivo del águila real. Cuanto mayor sea la media anual de LST, menor será la probabilidad de éxito reproductivo A pesar de la preferencia de esta población por áreas con influencia climática mediterránea (es decir, con temperaturas de verano más altas que las áreas más atlánticas), se encontró que la supervivencia de la cría del águila real en nidos expuestos disminuyó a medida que aumentaba el número de días con temperatura máxima de al menos 32,2 C. aumentó (Steenhof et al. 1997; Kochert et al. 2019). Nuestros resultados confirman que la temperatura de la superficie puede ser un factor limitante para el rendimiento reproductivo del águila real.
Se encontró que la media anual de la serie de tiempo EVI, un sustituto de la productividad de la vegetación, influye negativamente en el desempeño reproductivo del águila real. Por lo tanto, cuanto menor sea la productividad de la vegetación, mayor será la probabilidad de éxito reproductivo. Aunque está ampliamente aceptado que tasas de productividad primaria más altas permiten una mayor disponibilidad de energía para la reproducción (Connell y Orians 1964; Brasil et al. 2019), las áreas con productividad primaria baja en los territorios de reproducción del águila real generalmente se asocian con hábitats abiertos con escasamente áreas con vegetación, que indirectamente podrían mostrar las áreas de caza cercanas a los sitios de reproducción. Las áreas de matorral abierto son seleccionadas positivamente por el águila real como hábitats de caza porque la estructura de la vegetación favorece la detección de presas y el éxito de la caza (Tapia et al. 2007; Lado y Tapia 2012). Mas carroña alimentada por el águila real es frecuente cuando la presa natural es escasa (Watson 2010), y esta rapaz habita en zonas montañosas de ganadería en libertad en la región de estudio (Tapia et al. 2007). De hecho, las áreas abiertas dominado por matorrales bajos y pastizales también representan el hábitat típico seleccionado por los más presas comunes en la Península Ibérica: conejo de monte europeo (Oryctolagus cuniculus), liebre ibérica (Lepus granatensis) y perdiz roja (Alectoris rufa) (Tapia y Domínguez 2007; Tapia et al. 2010, 2014; Watson 2010). Investigaciones anteriores ya han demostrado que la la dinámica interanual de la productividad primaria afecta los patrones de colonización y ocupación del hábitat dinámica de los consumidores primarios y secundarios (Requena-Mullor et al. 2014; Fernández et al. 2016). Cambios interanuales en productividad primaria impulsada, por ejemplo, por fluctuaciones climáticas o procesos de sucesión natural así como los cambios abruptos causados por los incendios forestales podrían por lo tanto, impactan significativamente el desempeño de la especie.
La fecha del albedo mínimo en la temporada de crecimiento, un descriptor de la radiación neta en las superficies terrestres, se asoció con una menor probabilidad de éxito reproductivo del águila real. Como se mencionó anteriormente, las águilas suelen seleccionar los salientes de los acantilados para anidar (López-López et al. 2007; Tapia et al. 2007). Los sitios de anidación generalmente se encuentran en terrenos accidentados a gran altura en suelos desnudos con cubiertas vegetales bajas. Estas áreas están expuestas a altas temperaturas superficiales y bajas tasas de productividad primaria, lo que se traduce en altos valores de albedo. Por lo tanto, se encontró que la menor probabilidad de éxito reproductivo está asociada con la fecha en que los valores de albedo son mínimos en la temporada de crecimiento (es decir, los valores mas altos de productividad primaria). El cambio de uso del suelo se reconoce como uno de los principales factores que impulsan la dinámica del albedo superficial (ver, por ejemplo, Zhai et al. 2015; Hu et al. 2019). Sin embargo, la perturbación del fuego, que implica un intercambio crítico de energía y materia entre la tierra y la atmósfera, también puede afectar el balance de energía de la superficie al reducir inicialmente el albedo de la superficie (Gatebe et al. 2014) antes de recuperarse (a diferentes velocidades dependiendo del tiempo transcurrido desde el incendio y después del incendio Lyons et al. 2008; Gatebe et al. 2014; Dintwe et al. 2017; Torres et al. 2018) a sus condiciones de albedo previas al incendio. De hecho, nuestro estudio señala una respuesta con un retraso de 3 años del águila real a perturbaciones del presupuesto de energía superficial causadas por incendios forestales durante el período de 17 años Estudios previos en la España mediterránea mostraron que el conejo de campo y otras especies de presas potenciales colonizaron progresivamente las áreas quemadas (Moreno y Villafuerte 1995; Tapia et al. 2010, 2014), donde sus la abundancia aumentó durante al menos 5 años después del incendio (Rollan y Real 2011).El efecto de los incendios forestales en el estado físico del águila real se puede explicar por su impacto conjunto en el variación espacio-temporal de LST, productividad de la vegetación y albedo.
En conclusión, el desempeño reproductivo de el águila real, un depredador ápice en la parte superior nivel trófico—está influenciado por el funcionamiento del ecosistema. remotamente descriptores funcionales del ecosistema detectados, como el desviación estándar de LST (como sustituto de la dinámica del calor estacional), media anual de EVI (sustituto de la productividad primaria) y la fecha de mínimo albedo en la temporada de crecimiento (sustituto de net radiación en la superficie terrestre) se encontró que eran factores relevantes que afectan su aptitud ecológica en una amplia zona de transición bioclimática en el sur de Europa. La perturbación del fuego también afectó la ecología. aptitud de este depredador del ápice al afectar potencialmente disponibilidad de hábitat de las especies en los niveles tróficos más bajos, con un efecto positivo limitado a los 3 años después fuego (es decir, una respuesta retardada en el tiempo a las interrupciones del presupuesto de energía de la superficie). Nuestro estudio confirma la utilidad potencial al considerar el asunto y flujos de energía entre la superficie terrestre y la atmósfera para explicar el éxito reproductivo de las especies en los niveles tróficos superiores. Aunque localmente factores medidos como la abundancia de presas y Se sabe que la ubicación geográfica del nido es buena predictores del éxito reproductivo, anual en el campo.
No es una produccion propia, la fuente es Raptors of the World (.org)
Te puede interesar


La extinción masiva de especies amenaza el planeta. Actualmente se están llevando a cabo conversaciones sobre un nuevo tratado internacional para conservar la biodiversidad. ¿Qué es exactamente lo que está en juego?

Un millón de plantas y animales están en peligro de extinción. Exploramos qué podemos hacer para preservar la valiosa biodiversidad de la Tierra


La naturaleza tiene una capacidad inmensa para recuperarse después de las perturbaciones (lo lleva haciendo desde que surgió la vida)
Lo más visto

El futuro argentino parece potenciado por hidrocarburos y ser la turbina de la economía del país. Sin embargo, hay casos en que un proyecto petróleo trajo consecuencias inesperadas a distintos países

Bosques: la importancia de generar conocimiento para su manejo y conservación
Marcos Bach
Arbolado23/03/2026El Día Internacional de los Bosques es una jornada conmemorativa que se celebra anualmente el primer 21 de marzo desde 2013, establecida por Asamblea General de las Naciones Unidas el 21 de diciembre de 2012

Hasta Naciones Unidas advierte: modificar la Ley de Glaciares pone en riesgo a comunidades y ciudades
Marcos Bach
Agua y Glaciares23/03/2026Naciones Unidas, constitucionalistas, organizaciones socioambientales y hasta voces del gubernamental Consejo Federal de Medio Ambiente (Cofema) expusieron numerosos argumentos para que no se modifique la Ley de Glaciares. Queda en evidencia que solo las multinacionales mineras, y sus gobernadores y legisladores aliados, priorizan la actividad extractiva por sobre el acceso de la población al agua.

¿Qué es el "Super el Niño"? un fenómeno que podría transformar el clima global y el régimen de lluvias en Sudamérica
Marcos Bach
Cambio Climático23/03/2026Entre medidados y fines de 2026 podría consolidarse el evento climático, con riesgo de temperaturas récord, eventos meteorológicos extremos y un giro decisivo en la dinámica de precipitaciones del Cono Sur

Del uranio en la tierra al diseño de reactores, Argentina tiene todo lo necesario para un ciclo soberano que la posiciona en la elite global